Cranial Cavity, Skull Base
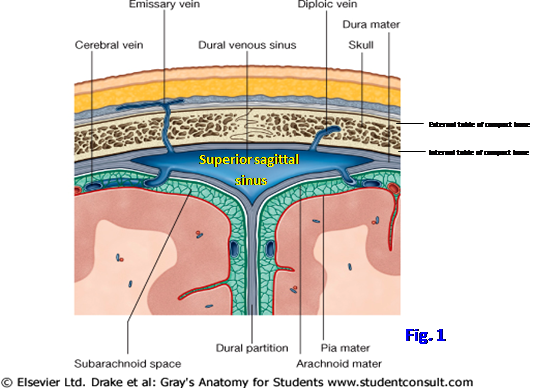
The skull consists of the cranium and the mandible (G11 7.2; G12 7.3; N6). The cranium is subdivided into the facial skeleton (viscerocranium) and the neural skeleton (neurocranium). The neurocranium is formed by the cranial vault and cranial base enclosing the cranial cavity, where the brain, meninges, and associated structures are housed. The roof of the cranial vault, or skull cap, is the calvaria (Note: many anatomists use the terms “cranial vault” and “calvaria” interchangeably). Most bones of the calvaria consist of internal and external tables of compact bone separated by cancellous bone (diploë) containing red bone marrow (Fig. 1). Venous channels (diploic veins) course through the diploë and have connections with scalp veins and dural venous sinuses (Fig. 1; G11 7.11D; G12 7.16D; N99, 101). The inner table of compact bone is thinner than the outer table and may be fractured by a blow that doesn’t fracture the outer table, which may make interpreting an x-ray difficult.
Some bones of the viscerocranium and neurocranium are joined by fibrous joints called sutures (G11 7.2, 7.3A-B, 7.6; G12 7.1, 7.3, 7.4A-B; N6, 8, 9). Where more than two bones of the cranial vault meet, the intervening fibrous connective tissue is called a fontanelle (G12 7.1A-D; N14). Sutures and fontanelles allow bones to overlap during passage through the birth canal, and the head of the newborn infant may be temporarily misshapen as a result (molding). Sutures and fontanelles also are important sites of growth in the skull because bone can grow only on edges (appositional growth), and growth of the cranial vault in the fetus, infant, and young child is necessary to accommodate rapid growth of the brain and meninges. Most cranial vault sutures begin to fuse (close) during the third decade of life, but remnants frequently indicate their location. Premature fusion of the fetus’ or infant’s sutures results in a skull deformity (craniosynostosis). For more information, see http://www.aafp.org/afp/20040615/2863.html . An important normal variation in sutures of the cranial vault is failure of the metopic (interfrontal) suture between the child’s right and left frontal bones to fuse, as it usually begins to do at two years of age. In about 8% of the population, the metopic suture doesn’t fuse and may be mistaken for a skull fracture in the x-ray of a trauma victim.
The coronal suture is located between the frontal bone and the two parietal bones and is important for longitudinal growth of the skull. Like the other cranial vault sutures, the coronal suture should appear largely fused in a middle-aged or older adult. The sagittal suture in the midline separates the two parietal bones and is important for growth in width of the skull. Bregma is the intersection of the coronal and sagittal sutures and indicates the location of the infant’s anterior fontanelle, the “soft spot” of a baby’s head, which is useful for assessing the infant’s degree of hydration, increased intracranial pressure, progress of ossification, etc.
The lambdoid suture is located posteriorly between the occipital bone and the two parietal bones and contributes to growth in length of the skull. The junction of the sagittal and lambdoid sutures is lambda, which indicates the site of the posterior fontanelle in the newborn infant. Laterally the squamosal suture joins the superior border of the squamous portion of the temporal bone with the parietal bone. Pterion is the “H”-shaped junction of the squamous part of the temporal bone, parietal bone, frontal bone, and greater wing of the sphenoid bone located in the floor of the temporal fossa. This gives an external indication of where the anterior branch of the middle meningeal artery forms a groove or canal in the internal surface of the cranial vault and may be torn by a skull fracture to produce an extradural (epidural) hematoma, which may be fatal within hours if untreated.
REMOVAL OF THE CALVARIA
1. Study the exposed part of the cranial vault. Attempt to identify remnants of the coronal suture between the frontal bone and the two parietal bones and the sagittal suturein the midline between the two parietal bones. These sutures are fused in middle-aged and older adults and may be completely obliterated. Bregma is the intersection of the coronal and sagittal sutures and indicates the location of the infant’s anterior fontanelle, the “soft spot” of a baby’s head.
2. Posteriorly look for the lambdoid suture between the two parietal bones and the occipital bone. The junction of the sagittal and lambdoid sutures is lambda, which indicates the site of the posterior fontanelle in the newborn infant. Laterally look for the squamosal suture between the superior border of the squamous portion of the temporal bone and the parietal bone. Identify pterion where the squamous part of the temporal bone, parietal bone, frontal bone, and greater wing of the sphenoid bone meet in an “H”-shaped junction.
3. Tie a string around the cranial vault so that anteriorly the string is located 1 cm above the supraorbital margin and posteriorly it is located 1 cm above the external occipital protuberance (inion) (Fig. 2). You may either use the string as your saw guide or use a pencil or indelible marker to draw a line along it. TRY TO AVOID CUTTING COMPLETELY THROUGH THE BONE OF THE CRANIAL VAULT. Following the string or mark, use a Stryker saw to cut PARTIALLY through the bone, into the diploë, around the circumference of the cranial vault (A⇒B). Then use a hammer and chisel to break the internal table of compact bone, leaving the dura mater intact. WHEN USING THE HAMMER AND CHISEL, DO NOT HIT THE BONE HARD AS THE BONE MAY BE VERY BRITTLE! Pry the calvaria away from the dura mater CAREFULLY and remove it. This may be difficult because of Sharpey’s fibers anchoring the bone to the periosteal layer of dura mater. These will appear as thin, hair-like structures protruding from the surface of the dura mater after the calvaria is removed.
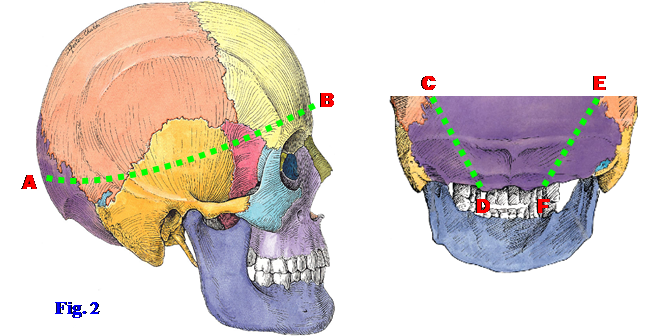
4. With the cadaver propped up in a sitting position or lying prone with a block under the chest so the neck can be flexed, scrape any remaining muscle from the bone in the occipital region down to the margin of the foramen magnum. Draw an oblique line on each side from where the cut edge of the skull intersects the lambdoid suture to the posterolateral edge of the foramen magnum (C⇒D and E⇒F). Saw through the outer part of the bone along those lines with a Stryker saw and use the mallet and chisel to free and gently remove this occipital wedge of bone. XXX
On the internal surface of the calvaria remnants of the coronal, sagittal, and lambdoid sutures often are visible. Locate a midline ridge of bone on the anterior part of the inner surface of the calvaria. This frontal crest gives attachment to a sagittal fold of dura mater that separates the cerebral hemispheres, the falx cerebri. The frontal crest terminates superiorly in the midline sulcus for the superior sagittal sinus (N9).
The superior sagittal sinus is located in the convex border of the falx cerebri, at its attachment to the calvaria (Fig. 3; G11 7.15A, 7.16A; G12 7.20A, 7.21A; N101, 102). The sinus becomes wider and deeper as it passes posteriorly and has lateral extensions, the lateral lacunae (N100). There typically are small, but sometimes deep, pits (arachnoid foveolae) in the bone of the sulcus for the superior sagittal sinus and in the shallow depressions made by the lateral lacunae. The arachnoid foveolae are impressions made by arachnoid granulations, clumps of arachnoid mater that project through the meningeal layer of dura mater into dural venous sinuses to return cerebrospinal fluid (CSF) from the subarachnoid space to venous blood (G11 7.13A-B, 7.14, 7.15A; G12 7.18A-B, 7.19, 7.20A; N100, 101, 108).
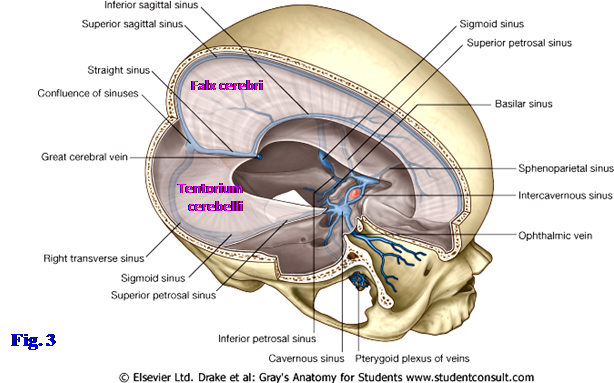
On the inner surface of the removed occipital wedge of bone the internal occipital protuberance should be obvious (G11 7.17; G12 7.6; N11). The superior sagittal sinus ends there in a confluence of sinuses or in continuity with the right transverse sinus (Fig. 6; G11 7.16, 7.18; G12 7.21, 7.22; N102, 103), which leaves a groove in the occipital bone at the posterior edge of the tentorium cerebelli. The left transverse sinus mainly receives blood from the straight sinus, which is located in the junction of the falx cerebri and tentorium cerebelli. The grooves for the transverse sinuses should be apparent in the occipital bone.
The meninges enclosing the brain consist of the same three layers as those around the spinal cord—dura mater, arachnoid mater, and pia mater—but some details of their arrangement are different (N101 vs. 163). The dura mater is the tough outer layer of meninges. The cranial dura mater differs from the spinal dura mater in that it consists of two layers, an outer periosteal layer and an inner meningeal layer. The periosteal layer of dura mater forms the periosteum on the inner surface of the cranial vault and is referred to as the “endocranium.” Therefore, it is not separated from the walls of the cranial cavity as the spinal dura mater is separated from the walls of the vertebral canal by the epidural space and its contents.
The meningeal layer of cranial dura mater is continuous with the spinal dura mater at the foramen magnum (G11 4.40, 7.20; G12 4.41, 7.20A, 7.24A; N102, 108). The meningeal layer and periosteal layer of cranial dura are fused except where they separate to form dural venous sinuses (G11 7.13A-B, 7.14; G12 7.18A-B, 7.19; N101, 103) and to form dural spaces for the hypophysis and for the trigeminal ganglia (trigeminal caves). Inward duplications of the meningeal layer form four dural folds (dural septa) that partly partition the cranial cavity into compartments for different parts of the brain (N102, 103). These compartments are important because increases in intracranial pressure and intracranial masses (e.g., tumors) can shift the soft brain against hard dural folds or even force part of the brain into another compartment (brain herniation) with potentially fatal consequences (see below). Alternatively, intracranial masses may result in compression of the cerebral vasculature with infarction of brain tissue.
The dural folds are the falx cerebri, tentorium cerebelli, diaphragma sellae, and falx cerebelli. The falx cerebri is a sickle-shaped, midsagittal meningeal fold that projects into the longitudinal fissure between the two cerebral hemispheres (Fig. 6; G11 7.15A, 7.16A; G12 7.20A, 7.21A; N101, 102). The superior sagittal sinus is located within the convex superior border of the falx cerebri, between the meningeal and periosteal layers of dura (Fig. 1, 3; N101, 102). The inferior sagittal sinus is located within the concave inferior border of the falx, just above the corpus callosum.
The falx cerebri fuses posteriorly with the superior surface of the tentorium cerebelli and partly divides the portion of the cranial cavity above the tentorium cerebelli (supratentorial compartment) into right and left compartments. An intracranial mass on one side of the supratentorial compartment (e.g., an extradural hematoma or tumor) may partially push the cerebral hemisphere of that side under the free inferior edge of the falx cerebri toward the contralateral side (subfalcine herniation), resulting in headache and possibly contralateral leg weakness.
The tentorium cerebelli is a fold of dura mater that separates the posterior portions of the cerebral hemispheres (occipital lobes) within the supratentorial compartment above from the cerebellum within the infratentorial compartment below (Fig. 3; G11 7.15A, 7.18; G12 7.20A, 7.22; N102, 103). It may be almost horizontal in orientation or angled posteroinferiorly. The tentorium is attached posteriorly to the margins of the grooves for the transverse sinuses on the occipital bone and laterally on each side to the apex of the petrous portion of the temporal bone at the margins of the shallow groove for the superior petrosal sinus (G11 7.15A, 7.17, 7.18, 7.23A; G12 7.6, 7.20A, 7.21, 7.22; N11, 103). Further anteriorly the tentorium attaches into the posterior and anterior clinoid processes (G11 7.18, 7.23A; G12 7.22, 7.27A; N11, 103). The tentorium cerebelli has a concave free edge anteromedially, forming an opening (tentorial notch or tentorial incisure) through which the midbrain descends from the supratentorial into the infratentorial compartment (G11 7.18 [right side], 7.21, 7.23A; G12 7.22, 7.25, 7.27A; N103, 105).
An increase in intracranial pressure or the development of a space-occupying lesion (e.g., a tumor) within the supratentorial compartment may cause partial herniation of the temporal lobe of the brain through the tentorial notch (transtentorial herniation). The temporal lobe may be damaged by the hard dura mater or may compress the ipsilateral oculomotor nerve and ipsilateral side of the midbrain (cerebral peduncle), producing ipsilateral pupillary dilation (and possibly abnormal extraocular movements) and contralateral hemiparesis (e.g., see http://rad.usuhs.mil/rad/herniation/ ). Parasympathetic nerve fibers in the oculomotor nerve are involved first in a compressive lesion because they are peripherally located within the nerve. A less frequent ascending transtentorial herniation may occur due to the mass effect of a posterior fossa tumor.
The falx cerebelli is a short, vertical, midline fold of dura mater between the two cerebellar hemispheres in the posterior cranial fossa (G11 7.15A, 7.16A; G12 7.20A, 7.21A; N 102, 142). It runs from the tentorium cerebelli above to the foramen magnum below. The small occipital sinus is located in its convex posterior border, which is attached to the internal occipital crest. The occipital sinus connects with the internal vertebral venous plexus within the vertebral canal, providing an ascending pathway for the spread of infection or cancer cells into the cranial cavity (G11 7.16A; G12 7.21A; N 102, 166) (e.g., see http://content.karger.com/produktedb/produkte.asp?typ=pdf&file=ENE2006056002136 ).
The diaphragma sellae is a circular, horizontal fold of meningeal dura mater that roofs the hypophyseal fossa and covers the pituitary gland (hypophysis). It has a circular opening for the passage of the infundibular (hypophyseal) stalk and hypophyseal vessels (G11 7.18, 7.22A-B; G12 7.22, 7.25, 7.26A-B; N103).
The cranial arachnoid mater is a filmy membrane comparable to the spinal arachnoid and is closely applied to the internal surface of the dura mater (G11 7.13A-B, 7.14; G12 7.18A-B, 7.19; N101). It therefore bridges the sulci of the cerebral hemispheres. There is no “subdural space’ between the dura and arachnoid mater as is sometimes described (and shown in N101)—the dura and arachnoid can be physically separated during life, but only by a pathological process (e.g., a subdural hematoma). Clumps of arachnoid villi (arachnoid granulations) protrude through the meningeal layer of dura mater into dural venous sinuses to return cerebrospinal fluid (CSF) to venous blood (G11 7.13A-B, 7.14, 7.15A; G12 7.18A-B, 7.19, 7.20A; N101, 108).
The pia mater is inseparable from the surface of the brain and faithfully follows the contours of the gyri and sulci (G11 7.13A-B; G12 7.18A-B; N101). The subarachnoid space between the arachnoid mater and pia mater is where CSF, which is produced by the choroid plexuses of the brain ventricles, circulates (N108). The development of the arachnoid and pia from the same embryonic layer of mesenchyme (leptomeninx) is indicated by thin connective tissue arachnoid trabeculae that bridge the subarachnoid space (G11 7.13A-B; G12 7.18A-B; N101). These trabeculae are thought to suspend the brain within the CSF-filled subarachnoid space. The presence of cerebral arteries on the surface of the brain means that a ruptured aneurysm there will result in bleeding into the subarachnoid space (subarachnoid hemorrhage). Aneurysms commonly involve the cerebral arterial circle (of Willis), usually the anterior part (G11 7.24, Table 7.5 [p. 624]; G12 7.28, Table 7.5 [p. 647]; N137-139) (e.g., see http://emedicine.medscape.com/article/1164341-overview#showall ).
CUTTING THE DURA-ARACHNOID MATER
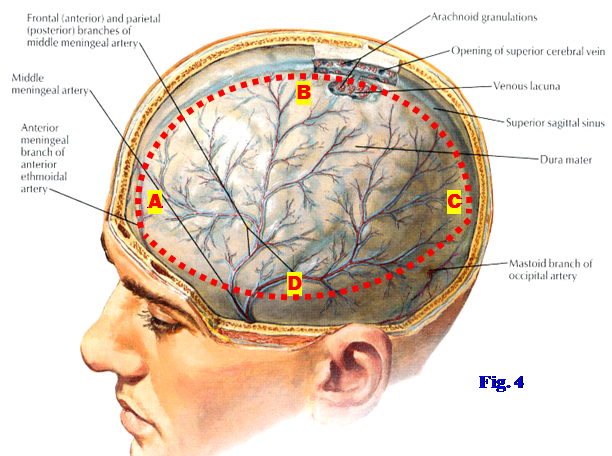
5. Identify the superior sagittal sinus within the convex superior border of the falx cerebri (Fig. 4). Make longitudinal incisions (using a pair of sharp scissors) through the dura mater approximately 1 cm lateral and parallel to, on both sides of, the superior sagittal sinus from the cut edge of the frontal bone to the cut edge of the occipital bone, where the calvaria was removed (Fig. 4, A⇒B⇒C). Make horizontal lateral incisions through the dura mater between the ends of the longitudinal cuts on each side parallel to the cut edge of the calvaria (A⇒D⇒C) and remove the mobilized dura mater. BE CAREFUL NOT TO CUT INTO THE BRAIN!
6. In order to remove the brain intact, the falx cerebri and tentorium cerebelli must be completely transected. Carefully lift the frontal lobes of the brain and cut the falx cerebri near its attachment to the crista galli. Pull the falx from between the cerebral hemispheres as far posteriorly as possible. The posterior inferior end of the falx cerebri will still be attached to the superior surface of the tentorium cerebelli.
7. Turning to the back of the head, gently lift the occipital lobe of the brain on one side. Using a scalpel (or curved Metzenbaum scissors, if available) sever the tentorium cerebelli under the occipital lobe laterally, parallel to the petrous portion of the temporal bone, from the tentorial notch to the transverse sinus (Fig. 5, A⇒B). Continue the cut medially along the transverse sinus to where the occipital wedge of bone was removed (B⇒C). At the occipital wedge cut downward through the transverse sinus and dura-arachnoid mater along the cut edge of bone. Repeat the same procedure on the other side. Before leaving the foramen magnum region, use the scalpel to make a transverse cut through the upper cervical portion of the spinal cord just inferior to foramen magnum. XXX
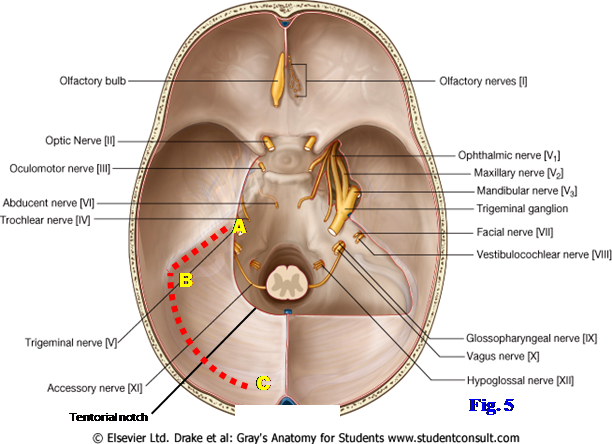
The twelve pairs of cranial nerves are the part of the peripheral nervous system attached to the brain. They are numbered in the rostral-to-caudal sequence that they pierce the dura mater in the floor of the cranial cavity (Fig. 5; G11 7.18, 7.19; G12 7.22, 7.23; N103, 117). The olfactory nerves (I) and optic nerves (II) are attached to the cerebrum. The olfactory bulbs and tracts and the optic nerves are actually brain tracts but are included among the cranial nerves. The true olfactory nerves are multiple thread-like filaments that traverse the cribriform plate of the ethmoid bone from the olfactory mucosa of the nasal cavity to synapse in the olfactory bulb (G11 Table 9.2 [p. 800]; G12 Table 9.2 [p. 818]; N42, 117, 118).
Each optic nerve is formed by the convergence of the axons of ganglion cells of the retina at the optic disc (optic papilla), which is the “blind spot” of the eye due to a lack of photoreceptor cells. The optic nerves traverse the optic canals from the orbits and unite at the optic chiasm, where there is a decussation (crossing) of fibers from the nasal halves of the two retinae (G11 Table 9.3 [p. 801]; G12 Table 9.3 [p. 819]; N 117, 119). The meninges and subarachnoid space extend around each optic nerve anteriorly as far as the eyeball (G11 7.39B-C; G12 7.35A-B; N87), so that increases in intracranial pressure (think increased CSF pressure) can inhibit venous return and result in swelling of the optic disc (papilledema). Papilledema is a contraindication to perform lumbar puncture, which may result in a fatal brain herniation, until imaging studies are done to rule out a mass lesion (see http://www.merck.com/mmpe/sec09/ch107/ch107e.html ).
Cranial nerves III-XII are attached to the brainstem, although cranial nerve XI (accessory) also has a spinal root arising from upper cervical spinal cord segments (G11 7.19; G12 7.23, 7.24A; N117). Three cranial nerves innervate the extraocular muscles that move the eye—the oculomotor (III), trochlear (IV), and abducent (abducens) (VI) nerves (G11 Table 9.4 [p. 804]; G12 Table 9.4 [p. 822]; N117, 120). All have a close relationship to the cavernous sinus, with III and IV located in its lateral wall and VI traversing it with the internal carotid artery (G11 7.22; G12 7.26; N103). Therefore, palsies of the extraocular muscles, often starting with loss of lateral movement of the eye by the lateral rectus muscle (innervated by VI), are common in cavernous sinus pathology. Nerves III, IV, and VI pass through the superior orbital fissure to reach the orbit from the middle cranial fossa.
The trigeminal nerve (V) is the great sensory nerve of the head but also has a motor component. Its small motor root innervates the muscles that develop from the first pharyngeal arch with special visceral efferent (SVE, branchial efferent, branchiomotor) nerve fibers. The large trigeminal ganglion (G11 7.18, 7.22A, Table 9.5 [p. 806]; G12 7.22, 7.26A, Table 9.5 [p. 824]; N103, 117, 121) contains the GSA nerve cell bodies of most sensory fibers within the three divisions of the trigeminal nerve. However, some GSA fibers are proprioceptive fibers, and the proprioceptive neuron cell bodies of V are the only first-order sensory neurons in the body with cell bodies located within the central nervous system (within the mesencephalic nucleus of the trigeminal nerve).
The three divisions of the trigeminal nerve leaving the ganglion are the ophthalmic division (V1) entering the orbit via the superior orbital fissure, the maxillary division (V2) entering the pterygopalatine fossa via the foramen rotundum, and the mandibular division (V3) entering the infratemporal fossa via the foramen ovale (Fig. 5;G11 7.18, 7.22A; G12 7.22, 7.26A; N13, 44, 45, 103). The small motor root also traverses the foramen ovale and joins the mandibular nerve just below the foramen (Fig. 5; G11 7.23A; G12 7.27A; N45). The mandibular division (V3)is the only division of the trigeminal nerve that contains motor fibers.
The ophthalmic, and usually the maxillary, divisions of the trigeminal are located within the lateral wall of the cavernous sinus (G11 7.22B-C; G12 7.26B-C; N103), so cavernous sinus thrombosis may result in increased (hyperesthesia) or decreased (hypoesthesia) sensation from their cutaneous distribution. The severe headache of cavernous sinus thrombosis may also be largely localized to the ophthalmic and maxillary distributions.
The facial (VII) and vestibulocochlear (VIII) nerves exit the posterior cranial fossa through the internal acoustic meatus (Fig. 5;G11 7.18, 7.20, 7.85E; G12 7.22, 7.24A, 7.74; N13, 97, 103, 123). The large motor root of the facial nerve carrying SVE fibers to muscles derived from the second pharyngeal arch is what is usually identified in dissection. Entering the internal acoustic meatus between the motor root of VII and the large vestibulocochlear nerve is the small nervus intermedius (intermediate nerve). The nervus intermedius carries SVA (taste) fibers from the anterior 2/3 of the tongue (via chorda tympani), GSA fibers from a small area of skin of the external ear canal, and GVE preganglionic parasympathetic fibers destined for the pterygopalatine and submandibular ganglia (G11 Table 9.9 [p. 813]; G12 Table 9.9; N122). The close physical relationship of VII and VIII mean that a Schwann cell tumor of the vestibulocochlear nerve (acoustic neuroma) may compress the facial nerve, but it can accommodate slow stretching, and ipsilateral facial paralysis rarely results. More commonly compression of the trigeminal nerve produces diminished sensation on the face (facial hypesthesia) and/or a decrease in the corneal reflex (mediated by cranial nerves V and VII) (see http://emedicine.medscape.com/article/882876-overview#showall ).
The glossopharyngeal (IX), vagus (X), and accessory (XI) nerves leave the medulla oblongata and exit the posterior cranial fossa via the jugular foramen (Fig. 5; G11 7.18, 7.20; G12 7.22, 7.24A; N13, 103, 124-126). The accessory nerve has cranial and spinal roots (N126). The cranial root of XI arises from the medulla oblongata and joins the vagus nerve, of which it is usually considered a part. SVE nerve fibers in X (some from the cranial root of XI) innervate skeletal muscles of the soft palate (except tensor veli palatini), pharynx (except stylopharyngeus), larynx, and upper half of the esophagus. The spinal root of XI arises from the upper five cervical spinal cord segments (G11 7.19, 7.20; G12 7.23, 7.24A). The rootlets join as they ascend, pass through the foramen magnum into the posterior cranial fossa, and exit it through the jugular foramen to supply GSE innervation to the sternocleidomastoid and trapezius muscles.
The shared exit of cranial nerves IX, X, and XI from the cranial cavity through the jugular foramen means that they can be injured together there (jugular foramen syndrome, e.g., http://emedicine.medscape.com/article/251009-overview#showall ) by a basal skull fracture, tumor, etc. The exact deficits depend on the extent to which each nerve is damaged, but commonly there is hoarseness, dysphagia, and ipsilateral sternocleidomastoid and trapezius paralysis and atrophy.
The rootlets of the hypoglossal nerve (XII) pass laterally from the medulla to reach the hypoglossal canal (Fig. 8; G11 7.18; G12 7.22; N103, 127). The rootlets join to form the hypoglossal nerve, which innervates the muscles of the tongue with GSE nerve fibers. The palatoglossus muscle is an exception innervated by X, but developmentally it is a muscle of the soft palate rather than the tongue.
Four of the cranial nerves leave the brain stem carrying preganglionic parasympathetic nerve fibers. They are cranial nerves III, VII, IX, and X (N161). Cranial nerve III (oculomotor nerve) carries preganglionic parasympathetic nerve fibers to the ciliary ganglion in the orbit (N120, 131, 161). Postganglionic parasympathetic fibers leave cell bodies within the ciliary ganglion to supply the sphincter pupillae and ciliary muscles of the eye.
Cranial nerve VII (facial nerve) carries preganglionic parasympathetic nerve fibers via its greater petrosal branch to the pterygopalatine ganglion and via its chorda tympani branch to the submandibular ganglion (N130, 132, 161). From cell bodies within the pterygopalatine ganglion, which is located within the pterygopalatine fossa, postganglionic parasympathetic fibers supply the lacrimal gland of the orbit and glands of the nasal cavity and palate. Postganglionic parasympathetic fibers from the submandibular ganglion in the floor of the oral cavity supply the submandibular and sublingual salivary glands and some smaller glands.
Cranial nerve IX (glossopharyngeal nerve) carries preganglionic parasympathetic fibers via its tympanic and lesser petrosal nerves to the otic ganglion within the infratemporal fossa (N124, 133, 161). Postganglionic fibers travel with the auriculotemporal nerve to supply the parotid gland.
Cranial nerve X (vagus nerve) carries preganglionic parasympathetic fibers to thoracic and abdominal viscera (N125, 161). Its distribution was studied with those regions.
REMOVING THE BRAIN
8. ATTEMPT TO REMOVE THE BRAIN INTACT. You may wish to get faculty help. Begin removing the brain from the front, studying the original position of the cranial nerves before they are cut. Gently elevate the frontal lobes from the floor of the anterior cranial fossa and displace the olfactory bulb and tractfrom the cribriform plate of the ethmoid bone on one side to remove with the brain (Fig. 5). Be aware that the actual olfactory nerves are multiple minute filaments that traverse the sieve-like cribriform plate of the ethmoid bone to reach the olfactory bulb from the olfactory mucosa of the nasal cavity.
9. All the subsequent cuts are best if a VERY SHARP scalpel or pair of scissors is used carefully.
10. Cut the olfactory tract that is to remain in the cranial cavity. Sever the optic nerves just posterior to the optic canals. This will be in front of the optic chiasm, which is to be removed with the brain. Cut the internal carotid arteries that ascend from the cavernous sinuses just medial to the anterior clinoid processes (G11 7.21, 7.22, 7.23; G12 7.25, 7.26, 7.27; N103, 135). If the stalk of the infundibulum is still intact entering the circular opening in the diaphragma sellae, cut it.
11. Gently lift the temporal lobes of the brain from the middle cranial fossa. Cut the oculomotor nerves just after they arise from the midbrain, before they enter the lateral walls of the cavernous sinuses. The delicate trochlear nerves may already be torn by this point. The trochlear nerves are the only cranial nerves that arise from the dorsal surface of the brainstem. If they are still intact, cut the trochlear nerves before they enter the free edges of the tentorium cerebelli en route to the lateral walls of the cavernous sinuses.
12. Cut each large trigeminal nerve as it passes anterolaterally from the pons to enter a separation between the periosteal and meningeal layers of dura mater overlying the apex of the petrous portion of the temporal bone, the trigeminal (Meckel’s) cave. The slender abducent nerves will probably already be torn by this time, but you should see them piercing the dura mater over the clivus in the posterior cranial fossa.
13. Sever thefacialand vestibulocochlear nerves just before they enter the internal acoustic meatus on each side. Continue lifting the brain posteriorly and transect the glossopharyngeal, vagus, and accessory nerves after they leave the medulla. They exit the posterior cranial fossa through the right and left jugular foramina.
14. If the vertebral arteries were not cut cleanly when the transverse cut was made through the foramen magnum to separate the brainstem and spinal cord, cut the arteries now. The vertebral arteries were previously seen on the posterior arch of the atlas within the suboccipital triangles. Note that the arteries ascend through the foramen magnum and join to form the basilar artery, which runs along the ventral surface of the pons in the midline. If the rootlets of the hypoglossal nerves are still intact as they pass from the medulla laterally into the hypoglossal canals, cut them.
15. Having severed the roots of the cranial nerves, the stalk of the pituitary gland, the internal carotid arteries, and the vertebral arteries, carefully remove the (hopefully) intact brain and set it in the provided bucket of preservative for later study. XXX
The meninges enclosing the brain consist of the same three layers as those around the spinal cord—dura mater, arachnoid mater, and pia mater—but some details of their arrangement are different (Ne 101vs. 163). The dura mater is the tough outer layer of meninges. The cranial dura mater differs from the spinal dura mater in that it consists of two layers, an outer periosteal layer and an inner meningeal layer. The periosteal layer of dura mater forms the periosteum on the inner surface of the cranial vault and is referred to as the “endocranium.” Therefore, it is not separated from the walls of the cranial cavity as the spinal dura mater is separated from the walls of the vertebral canal by the epidural space and its contents.
The meningeal layer of cranial dura mater is continuous with the spinal dura mater at the foramen magnum (Gr 4.40, 7.20; N102, 108). The meningeal layer and periosteal layer of cranial dura are fused except where they separate to form dural venous sinuses (Gr 7.13A-B, 7.14; N101, 103) and to form dural spaces for the hypophysis and for the trigeminal ganglia (trigeminal caves). Inward duplications of the meningeal layer form four dural folds (dural septa) that partly partition the cranial cavity into compartments for different parts of the brain (N102, 103). These compartments are important because increases in intracranial pressure and intracranial masses (e.g., tumors) can shift the soft brain against hard dural folds or even force part of the brain into another compartment (brain herniation) with potentially fatal consequences (see below). Alternatively, intracranial masses may result in compression of the cerebral vasculature with infarction of brain tissue.
The dural folds are the falx cerebri, tentorium cerebelli, diaphragma sellae, and falx cerebelli. The falx cerebri is a sickle-shaped, midsagittal meningeal fold that projects into the longitudinal fissure between the two cerebral hemispheres (Fig. 6; Gr 7.15A, 7.16A; N101, 102). The superior sagittal sinus is located in the convex superior border of the falx cerebri, between the meningeal and periosteal layers of dura (Fig. 2, 6; N101, 102). The inferior sagittal sinus is located in the concave inferior border of the falx, just above the corpus callosum.
The falx cerebri fuses posteriorly with the superior surface of the tentorium cerebelli and partly divides the portion of the cranial cavity above the tentorium cerebelli (supratentorial compartment) into right and left compartments. An intracranial mass on one side of the supratentorial compartment (e.g., an extradural hematoma or tumor) may partially push the cerebral hemisphere of that side under the free inferior edge of the falx cerebri toward the contralateral side (subfalcine herniation), resulting in headache and possibly contralateral leg weakness.
The tentorium cerebelli is a fold of dura mater that separates the posterior portions of the cerebral hemispheres (occipital lobes) in the supratentorial compartment above from the cerebellum in the infratentorial compartment below (Fig. 6; Gr 7.15A, 7.18; N102, 103). It may be almost horizontal in orientation or angled posteroinferiorly. The tentorium is attached posteriorly to the margins of the grooves for the transverse sinuses on the occipital bone and laterally on each side to the apex of the petrous portion of the temporal bone at the margins of the shallow groove for the superior petrosal sinus (Gr 7.15A, 7.17, 7.18, 7.23A; N11, 103). Further anteriorly the tentorium attaches into the posterior and anterior clinoid processes (Gr 7.18, 7.23A; N11, 103). The tentorium cerebelli has a concave free edge anteromedially, forming an opening (tentorial notch or tentorial incisure) through which the midbrain descends from the supratentorial into the infratentorial compartment (Gr 7.18 right side, 7.21, 7.23A; N103, 105).
An increase in intracranial pressure or the development of a space-occupying lesion (e.g., a tumor) in the supratentorial compartment may cause partial herniation of the temporal lobe of the brain through the tentorial notch (transtentorial herniation). The temporal lobe may be damaged by the hard dura mater or may compress the ipsilateral oculomotor nerve and ipsilateral side of the midbrain (cerebral peduncle), producing ipsilateral pupillary dilation (and possibly abnormal extraocular movements) and contralateral hemiparesis (e.g., see http://rad.usuhs.mil/rad/herniation/ ). Parasympathetic nerve fibers in the oculomotor nerve are involved first in a compressive lesion because they are peripherally located within the nerve. A less frequent ascending transtentorial herniation may occur due to the mass effect of a posterior fossa tumor.
The falx cerebelli is a short, vertical, midline fold of dura mater between the two cerebellar hemispheres in the posterior cranial fossa (Gr 7.15A, 7.16A; N102, 142). It runs from the tentorium cerebelli above to the foramen magnum below. The small occipital sinus is located in its convex posterior border, which is attached to the internal occipital crest. The occipital sinus connects with the internal vertebral venous plexus in the vertebral canal, providing an ascending pathway for the spread of infection or cancer cells into the cranial cavity (Gr 7.16A; N102, 166) (e.g., see http://content.karger.com/produktedb/produkte.asp?typ=pdf&file=ENE2006056002136 ).
The diaphragma sellae is a circular, horizontal fold of meningeal dura mater that roofs the hypophyseal fossa and covers the pituitary gland (hypophysis). It has a circular opening for the passage of the infundibular (hypophyseal) stalk and hypophyseal vessels (Gr 7.18, 7.22A-B; N103).
The cranial arachnoid mater is a filmy membrane comparable to the spinal arachnoid and is closely applied to the internal surface of the dura (Gr 7.13A-B, 7.14; N101). It therefore bridges the sulci of the cerebral hemispheres. There is no “subdural space’ between the dura and arachnoid mater as is sometimes described (and shown in N101)—the dura and arachnoid can be physically separated, but only by a pathological process (e.g., a subdural hematoma). Clumps of arachnoid villi (arachnoid granulations) protrude through the meningeal layer of dura mater into dural venous sinuses to return cerebrospinal fluid (CSF) to venous blood (Gr 7.13A-B, 7.14, 7.15A; N101, 108).
The pia mater is inseparable from the surface of the brain and faithfully follows the contours of the gyri and sulci (Gr 7.13A-B; N101). The subarachnoid space between the arachnoid mater and pia mater is where CSF, which is produced in the choroid plexus of the brain ventricles, circulates (N108). The development of the arachnoid and pia from the same embryonic layer of mesenchyme (leptomeninx) is indicated by thin connective tissue arachnoid trabeculae that bridge the subarachnoid space (Gr 7.13A-B; N101). These trabeculae are thought to suspend the brain in the CSF-filled subarachnoid space. The presence of cerebral arteries on the surface of the brain means that a ruptured aneurysm there will result in bleeding into the subarachnoid space (subarachnoid hemorrhage). Aneurysms commonly involve the cerebral arterial circle (of Willis), usually the anterior part (Gr 7.24, Table 7.5 [p. 624]; N137-139) (e.g., see http://emedicine.medscape.com/article/1164341-overview).
CUTTING THE DURA-ARACHNOID MATER
7. Identify the superior sagittal sinus in the convex superior border of the falx cerebri (Fig. 7). Make longitudinal incisions (using a pair of sharp scissors) through the dura mater approximately 1 cm lateral and parallel to, on both sides of, the superior sagittal sinus from the cut edge of the frontal bone to the cut edge of the occipital bone, where the calvaria was removed (Fig. 7, A⇒B⇒C). Make horizontal lateral incisions through the dura mater between the ends of the longitudinal cuts on each side parallel to the cut edge of the calvaria (A⇒D⇒C) and remove the mobilized dura mater. BE CAREFUL TO NOT CUT INTO THE BRAIN!
8. In order to remove the brain intact, the falx cerebri and tentorium cerebelli must be completely transected. Carefully lift the frontal lobes of the brain and cut the falx cerebri near its attachment to the crista galli. Pull the falx from between the cerebral hemispheres as far posteriorly as possible. The posterior inferior end of the falx cerebri will still be attached to the superior surface of the tentorium cerebelli.
9. Turning to the back of the head, gently lift the occipital lobe of the brain on one side. Using a scalpel (or curved Metzenbaum scissors, if available) sever the tentorium cerebelli under the occipital lobe laterally, parallel to the petrous portion of the temporal bone, from the tentorial notch to the transverse sinus (Fig. 8, A⇒B). Continue the cut medially along the transverse sinus to where the occipital wedge of bone was removed (B⇒C). At the occipital wedge cut downward through the transverse sinus and dura-arachnoid mater along the cut edge of bone. Repeat the same procedure on the other side. Before leaving the foramen magnum region, use the scalpel to make a transverse cut through the upper cervical portion of the spinal cord just inferior to foramen magnum. XXX
The twelve pairs of cranial nerves are the part of the peripheral nervous system attached to the brain. They are numbered in the rostral-to-caudal sequence that they pierce the dura mater in the floor of the cranial cavity (Fig. 8; Gr 7.18, 7.19; N103, 117). The olfactory nerves (I) and optic nerves (II) are attached to the cerebrum. The olfactory bulbs and tracts and the optic nerves are actually brain tracts but are included among the cranial nerves. The true olfactory nerves are multiple thread-like filaments that traverse the cribriform plate of the ethmoid bone from the olfactory mucosa of the nasal cavity to synapse in the olfactory bulb (Gr Table 9.2 [p. 800]; N42, 117, 118).
Each optic nerve is formed by the convergence of the axons of ganglion cells of the retina at the optic disc (optic papilla), which is the “blind spot” of the eye due to a lack of photoreceptor cells. The optic nerves traverse the optic canals from the orbits and unite at the optic chiasm, where there is a decussation (crossing) of fibers from the nasal halves of the two retinae (Gr Table 9.3 [p. 801]; N117, 119). The meninges and subarachnoid space extend around each optic nerve anteriorly as far as the eyeball (Gr 7.39B-C; N87), so that increases in intracranial pressure (think increased CSF pressure) can inhibit venous return and result in swelling of the optic disc (papilledema). Therefore, papilledema is a contraindication to perform lumbar puncture, which may result in a fatal brain herniation, until imaging studies are done to rule out a mass lesion (see http://www.merck.com/mmpe/sec09/ch107/ch107e.html ).
Cranial nerves III-XII are attached to the brainstem, although cranial nerve XI (accessory) also has a spinal root arising from upper cervical spinal cord segments (Gr 7.19; N117). Three cranial nerves innervate the extraocular muscles that move the eye—the oculomotor (III), trochlear (IV), and abducent (abducens) (VI) nerves (Gr Table 9.4 [p. 804]; N117, 120). All have a close relationship to the cavernous sinus, with III and IV located in its lateral wall and VI traversing it with the internal carotid artery (Gr 7.22; N103). Therefore, palsies of the extraocular muscles, often starting with loss of lateral movement of the eye by the lateral rectus muscle (innervated by VI), are common in cavernous sinus pathology. Nerves III, IV, and VI pass through the superior orbital fissure to reach the orbit from the middle cranial fossa.
The trigeminal nerve (V) is the great sensory nerve of the head but also has a motor component. Its small motor root innervates the muscles that develop from the first pharyngeal arch with special visceral efferent (SVE, branchial efferent, branchiomotor) nerve fibers. The large trigeminal ganglion (Gr 7.18, 7.22A, Table 9.5 [p. 806]; N103, 117, 121) contains the GSA nerve cell bodies of most sensory fibers within the three divisions of the trigeminal nerve. However, some GSA fibers are proprioceptive fibers, and the proprioceptive neuron cell bodies of V are the only first-order sensory neurons in the body with cell bodies located within the central nervous system (within the mesencephalic nucleus of the trigeminal nerve).
The three divisions of the trigeminal nerve leaving the ganglion are the ophthalmic division (V1) entering the orbit via the superior orbital fissure, the maxillary division (V2) entering the pterygopalatine fossa via the foramen rotundum, and the mandibular division (V3) entering the infratemporal fossa via the foramen ovale (Fig. 8;Gr 7.18, 7.22A; N13, 44, 45, 103). The small motor root also traverses the foramen ovale and joins the mandibular nerve just below the foramen (Fig. 7.23A; N45). The mandibular division (V3) is the only division of the trigeminal nerve that contains motor fibers.
The ophthalmic, and usually the maxillary, divisions of the trigeminal are located in the lateral wall of the cavernous sinus (Gr 7.22B-C; N103), so cavernous sinus thrombosis may result in increased (hyperesthesia) or decreased (hypoesthesia) sensation from their cutaneous distribution. The severe headache of cavernous sinus thrombosis may also be largely localized to the ophthalmic and maxillary distributions.
The facial (VII) and vestibulocochlear (VIII) nerves exit the posterior cranial fossa through the internal acoustic meatus (Fig. 8; Gr 7.18, 7.20, 7.85E; N13, 97, 103, 123). The large motor root of the facial nerve carrying SVE fibers to muscles derived from the second pharyngeal arch is what is usually identified in dissection. Entering the internal acoustic meatus between the motor root of VII and the large vestibulocochlear nerve is the small nervus intermedius (intermediate nerve). The nervus intermedius carries SVA (taste) fibers from the anterior 2/3 of the tongue (via chorda tympani), GSA fibers from a small area of skin of the external ear canal, and GVE preganglionic parasympathetic fibers destined for the pterygopalatine and submandibular ganglia (Gr Table 9.9 [p. 813]; N122). The close physical relationship of VII and VIII mean that a Schwann cell tumor of the vestibulocochlear nerve (acoustic neuroma) may compress the facial nerve, but it can accommodate slow stretching, and ipsilateral facial paralysis rarely results. More commonly compression of the trigeminal nerve produces diminished sensation on the face (facial hypesthesia) and/or a decrease in the corneal reflex (mediated by cranial nerves V and VII) (see http://emedicine.medscape.com/article/882876-overview ).
The glossopharyngeal (IX), vagus (X), and accessory (XI) nerves leave the medulla oblongata and exit the posterior cranial fossa via the jugular foramen (Fig. 8; Gr 7.18, 7.20; N13, 103, 124-126). The accessory nerve has cranial and spinal roots (N126). The cranial root of XI arises from the medulla oblongata and joins the vagus nerve, of which it is usually considered a part. SVE nerve fibers in X (some from the cranial root of XI) innervate skeletal muscles of the soft palate (except tensor veli palatini), pharynx (except stylopharyngeus), larynx, and upper half of the esophagus. The spinal root of XI arises from the upper five cervical spinal cord segments (Gr 7.19, 7.20). The rootlets join as they ascend, pass through the foramen magnum into the posterior cranial fossa, and exit it through the jugular foramen to supply GSE innervation to the sternocleidomastoid and trapezius muscles.
The shared exit of cranial nerves IX, X, and XI from the cranial cavity through the jugular foramen means that they can be injured together there (jugular foramen syndrome, e.g., http://emedicine.medscape.com/article/251009-overview ) by a basal skull fracture, tumor, etc. The exact deficits depend on the extent to which each nerve is damaged, but commonly there is hoarseness, dysphagia, and ipsilateral sternocleidomastoid and trapezius paralysis and atrophy.
The rootlets of the hypoglossal nerve (XII) pass laterally from the medulla to reach the hypoglossal canal (Fig. 8; Gr 7.18; N103, 127). The rootlets join to form the hypoglossal nerve, which innervates the muscles of the tongue with GSE nerve fibers. The palatoglossus muscle is an exception innervated by X, but developmentally it is a muscle of the soft palate rather than the tongue.
Four of the cranial nerves leave the brain stem carrying preganglionic parasympathetic nerve fibers. They are cranial nerves III, VII, IX, and X (N161). Cranial nerve III (oculomotor nerve) carries preganglionic parasympathetic nerve fibers to the ciliary ganglion in the orbit (N120, 131, 161). Postganglionic parasympathetic fibers leave cell bodies in the ciliary ganglion to supply the sphincter pupillae and ciliary muscles of the eye.
Cranial nerve VII (facial nerve) carries preganglionic parasympathetic nerve fibers via its greater petrosal branch to the pterygopalatine ganglion and via its chorda tympani branch to the submandibular ganglion (N130, 132, 161). From cell bodies in the pterygopalatine ganglion, which is located in the pterygopalatine fossa, postganglionic parasympathetic fibers supply the lacrimal gland of the orbit and glands of the nasal cavity and palate. Postganglionic parasympathetic fibers from the submandibular ganglion in the floor of the oral cavity supply the submandibular and sublingual salivary glands and some smaller glands.
Cranial nerve IX (glossopharyngeal nerve) carries preganglionic parasympathetic fibers via its tympanic and lesser petrosal nerves to the otic ganglion in the infratemporal fossa (N124, 133, 161). Postganglionic fibers travel with the auriculotemporal nerve to supply the parotid gland.
Cranial nerve X (vagus nerve) carries preganglionic parasympathetic fibers to thoracic and abdominal viscera (N125, 161). Its distribution was studied with those regions.
REMOVING THE BRAIN
10. ATTEMPT TO REMOVE THE BRAIN INTACT. It might be wise to get faculty help. Begin removing the brain from the front, studying the original position of the cranial nerves before they are cut. Gently elevate the frontal lobes from the floor of the anterior cranial fossa and displace the olfactory bulb and tractfrom the cribriform plate of the ethmoid bone on one side to remove with the brain (Fig. 8). Be aware that the actual olfactory nerves are multiple minute filaments that traverse the sieve-like cribriform plate of the ethmoid bone to reach the olfactory bulb from the olfactory mucosa of the nasal cavity.
11. All the subsequent cuts are best if a VERY SHARP scalpel or pair of scissors is used carefully.
12. Cut the olfactory tract that is to be left in the cranial cavity. Sever the optic nerves just posterior to the optic canals. This will be in front of the optic chiasm, which is to be removed with the brain. Cut the internal carotid arteries that ascend from the cavernous sinuses just medial to the anterior clinoid processes (Gr 7.21, 7.22, 7.23; N103, 135). If the stalk of the infundibulum is still intact entering the circular opening in the diaphragma sellae, cut it.
13. Gently lift the temporal lobes of the brain from the middle cranial fossa. Cut the oculomotor nerves just after they arise from the midbrain, before they enter the lateral walls of the cavernous sinuses. The delicate trochlear nerves may already be torn by this point. The trochlear nerves are the only cranial nerves that arise from the dorsal surface of the brainstem. If they are still intact, cut the trochlear nerves before they enter the free edges of the tentorium cerebelli en route to the lateral walls of the cavernous sinuses.
14. Cut each large trigeminal nerve as it passes anterolaterally from the pons to enter a separation between the periosteal and meningeal layers of dura mater overlying the apex of the petrous portion of the temporal bone, the trigeminal (Meckel’s) cave. The slender abducent nerves will probably already be torn by this time, but you should see them piercing the dura mater over the clivus in the posterior cranial fossa.
15. Sever the facial and vestibulocochlear nerves just before they enter the internal acoustic meatus on each side. Continue lifting the brain posteriorly and transect the glossopharyngeal, vagus, and accessory nerves after they leave the medulla. They exit the posterior cranial fossa through the right and left jugular foramina.
16. If the vertebral arteries were not cut cleanly when the transverse cut was made through the foramen magnum to separate the brainstem and spinal cord, cut the arteries now. The vertebral arteries were previously seen on the posterior arch of the atlas in the suboccipital triangles. Note that they ascend through the foramen magnum and join to form the basilar artery, which runs along the ventral surface of the pons in the midline. If the rootlets of the hypoglossal nerves are still intact as they pass from the medulla laterally into the hypoglossal canals, cut them.
17. Having severed the roots of the cranial nerves, the stalk of the pituitary gland, the internal carotid arteries, and the vertebral arteries, carefully remove the (hopefully) intact brain and set it in the provided bucket of preservative for later study. XXX
The floor of the cranial cavity is divided into anterior, middle, and posterior cranial fossae (Gr 7.17; N8, 11). The shallow anterior cranial fossa is formed anterolaterally by the orbital parts of the frontal bone, in the midline by the ethmoid bone, and posteriorly by the anterior part of the body and the lesser wings of the sphenoid bone. The anterior cranial fossa is related below to the nasal cavities in the midline and the orbits laterally. The anterior cranial fossa supports the frontal lobes of the brain.
The olfactory bulb overlies the cribriform plate of the ethmoid beside the crista galli (Fig. 8; Gr 7.18; N13, 118). Passing posteriorly toward the cerebrum from the olfactory bulb is the olfactory tract.
The middle cranial fossa is formed by parts of the sphenoid and temporal bones (Fig. 9; Gr 7.17; N8, 11). It is separated from the anterior cranial fossa by the anterior edge of the chiasmatic sulcus (prechiasmatic groove) in the midline and the posterior edges of the lesser wings of the sphenoid laterally (Note: some authors include the chiasmatic sulcus in the anterior cranial fossa). The posterior boundaries of the middle cranial fossa are the dorsum sellae in the midline and the apices of the petrous portions of the temporal bones on each side. The middle cranial fossa laterally supports the temporal lobes of the brain.
The body of the sphenoid bone in the middle cranial fossa forms the sella turcica. It consists of an anterior wall (tuberculum sellae), deep central depression for the pituitary gland (hypophyseal fossa), and a posterior wall (dorsum sellae) (Gr 7.17B; N8, 11). The sella turcica often has a close relationship inferiorly to the sphenoid sinuses, depending on the extent to which the sinuses invade the bone (Gr 7.66; N8, 38, 105), and this is clinically important in sinus infections and in the neurosurgical excision of a pituitary tumor via a nasal cavity approach (transsphenoidal hypophysectomy) (e.g., see http://www.e-mjm.org/2006/v61n3/Transseptal_Transphenoidal_Hypophysectomy.pdf ). A tumor of the pituitary gland or chronic increased intracranial pressure may cause radiographic changes in the sella turcica. To each side of the sella turcica the floor of the middle cranial fossa is formed by a greater wing of the sphenoid bone (Gr 7.17B; N11). The more posterior and lateral part of the floor of the middle cranial fossa is formed by the petrous portion of the temporal bone.
The middle cranial fossa contains a number of important structures. The optic nerve enters the optic canal. The small, but important, ophthalmic artery branches from the internal carotid artery to enter the optic canal inferior to the optic nerve (N85, 141). The diaphragma sellae covers the pituitary gland in the hypophyseal fossa. A superior expansion of a pituitary tumor may compress the optic chiasm to produce visual disturbances.
Just lateral to the sella turcica on each side is the cavernous sinus. This dural venous sinus has important venous connections with the facial vein, pterygoid venous plexus, and cerebral veins (Gr 7.16, 7.22B-C; N85, 103). It also has important spatial relationships to other structures. The superior and inferior ophthalmic veins connect it to the facial vein, providing a pathway for the spread of infection from the middle 1/3 of the face and the orbit (N70, 85). The inferior ophthalmic vein may drain into the superior ophthalmic vein proximal to the sinus. A sphenoidal emissary vein connects the cavernous sinus with the pterygoid plexus of veins in the infratemporal fossa. This vein may traverse either the foramen ovale or an occasional foramen, the sphenoidal emissary foramen (of Vesalius) (N13).
The right and left cavernous sinuses are connected across the sella turcica by intercavernous sinuses (Fig. 6; N103). Therefore, one of the usual indications of a cavernous sinus thrombosis that involves structures in one orbit is that the signs and symptoms usually spread to the other orbit within 24-48 hours (see http://emedicine.medscape.com/article/791704-overview).
The cavernous sinus is so named because it is partly subdivided by connective tissue septa into smaller communicating spaces, so it resembles cavernous (erectile) tissue (N103). This slows the flow of blood through the sinus and provides a large endothelial surface area for thrombus formation due to infection. Passing through the sinus to further interfere with the flow of blood are the internal carotid artery and abducent nerve. The artery is surrounded by the internal carotid plexus of sympathetic nerve fibers (N131). Both the internal carotid artery and abducent nerve are normally separated from the sinus’ blood by an endothelial layer.
The internal carotid artery enters the cavernous sinus from the carotid canal above the foramen lacerum and courses forward in the carotid groove beside the sella turcica. Near the anterior end of the cavernous sinus, the artery curves upward and backward, emerging from the cavernous sinus medial to the anterior clinoid process. The curved part of the artery where it leaves the cavernous sinus is referred to by radiologists as the “anterior loop of the carotid siphon” (Gr 7.17B, 7.18, 7.22, 7.23, 7.25B; N103, 135, 141). The anterior loop of the carotid siphon is why coronal sections of the anterior part of the cavernous sinus show the artery cut twice. The small ophthalmic artery branches from the internal carotid artery just after it emerges from the cavernous sinus. The artery enters the optic canal below the optic nerve.
A fracture of the cranial base may tear the internal carotid artery in the cavernous sinus or an aneurysm may rupture there, creating an arteriovenous fistula. The higher pressure arterial blood forces retrograde blood flow through the ophthalmic veins, causing a pulsating protrusion of the eyeball (pulsating exophthalmos) and swelling of the conjunctiva lining the eyelids and anterior surface of the eye (chemosis). The cranial nerves associated with the sinus also may be affected.
The abducent nerve (VI) traverses the cavernous sinus adjacent to the internal carotid artery (Gr 7.22B-C, 7.23; N103) and often is the first nerve affected in a cavernous sinus thrombosis. This would result in the patient having difficulty moving the eye to look laterally on the affected side. The oculomotor (III), trochlear (IV), ophthalmic division of the trigeminal (V1), and often the maxillary division (V2) are embedded in the lateral wall of the sinus. These nerves may be damaged in a cavernous sinus infection or by a pituitary tumor that expands laterally.
The trigeminal (Meckel’s) cave housing the trigeminal ganglion shares part of its medial wall with the cavernous sinus (Gr 7.18, 7.22; N103). The superior orbital fissure, foramen rotundum, foramen ovale, and foramen spinosum form a crescent of foramina lateral to the sella turcica (Gr 7.17; N13), and divisions of the trigeminal nerve leave the ganglion to exit the cranial cavity through the first three of these openings (N44). The ophthalmic division (V1) enters the orbit through the superior orbital fissure. The maxillary division (V2) enters the pterygopalatine fossa through the foramen rotundum. The mandibular division (V3) enters the infratemporal fossa through the foramen ovale. The small motor root of the trigeminal nerve runs below the ganglion and leaves the middle cranial fossa with V3, which it joins just below the foramen ovale (N45).
The middle meningeal branch of the maxillary artery enters the cranial cavity through the foramen spinosum. It passes laterally and divides into anterior (frontal) and posterior (parietal) branches that ascend on the lateral wall of the cranial vault in the temporoparietal region (Gr 7.17, 7.18; N8, 100, 103). The arteries usually leave easily identifiable grooves in the bone. The anterior branch of the middle meningeal artery typically occupies a deep groove or an actual canal in the region of pterion (N6). Therefore, a fracture of the skull at pterion may tear the artery, and the escaping blood separates the dura mater from the bone (epidural hematoma or extradural hematoma) (Gr 7.13C). An epidural hematoma may be fatal within hours if undetected and untreated, although the clinical course is variable (e.g., http://www.emedicine.com/neuro/topic574.htm ). Epidural hematomas have a characteristic biconvex appearance on diagnostic images (Fig. 10).
The superior and inferior petrosal sinuses drain the cavernous sinus posteriorly. The superior petrosal sinus lies in the anterolateral attached margin of the tentorium cerebelli along the apex of the petrous temporal bone. It drains blood from the cavernous sinus to the junction of the transverse and sigmoid sinuses (Fig. 6; Gr 7.16, 7.18; N102, 103).
The inferior petrosal sinus drains more inferiorly, along the petrooccipital fissure in the posterior cranial fossa. It passes through the jugular foramen with the sigmoid sinus to end in the internal jugular vein. The two inferior petrosal sinuses are connected by the basilar plexus (basilar sinus) on the clivus (Fig. 6; Gr 7.16; N102, 103). The basilar plexus communicates through foramen magnum with the internal vertebral venous plexus, providing a pathway (along with the occipital sinus) for the spread of cancer cells (e.g., from a thoracic, abdominal, or pelvic tumor) or infections into the cranial cavity.
18. In order to study many of the structures below, you will need to gently remove (strip) the dura mater from the subjacent bone. PLEASE BE CAREFUL WHEN YOU DO THIS OR YOU MAY DAMAGE OR REMOVE THE STRUCTURES TO BE IDENTIFIED!
19. Identify the groove over the cribriform plate occupied by the remaining olfactory bulb and follow the olfactory tract to its cut end. Find the cut ends of the optic nerves emerging from the optic canals. Lift the end of an optic nerve to expose the ophthalmic artery entering the optic canal after branching from the internal carotid artery. Note that the internal carotid artery ascends from the cavernous sinus medial to the anterior clinoid process.
20. Look for the delicate infundibular stalk, which was cut or torn emerging through the diaphragma sellae from the hypophyseal fossa. Carefully incise the roof of the cavernous sinus on one side. Find the internal carotid artery passing forward through the cavernous sinus with the slender abducent nerve. In the lateral wall of the sinus clean the oculomotor, trochlear, and ophthalmic nerves as they travel toward the superior orbital fissure.
21. Incise the dural roof of the trigeminal cave to expose the trigeminal ganglion. The ophthalmic division has already been exposed leaving the ganglion. Now find the maxillary division of the trigeminal nerve exiting the middle cranial fossa through the foramen rotundum. Is the maxillary division embedded in the lateral wall of the cavernous sinus in this cadaver? Clean the mandibular division leaving through the foramen ovale and note that it is lateral to the cavernous sinus. The delicate lesser petrosal nerve (IX) also may be visible approaching the foramen ovale from the side (Gr 7.22A). We will discuss how it arrives there later.
22. Clean the middle meningeal artery as it enters the cranial cavity from the infratemporal fossa through foramen spinosum. Trace the artery laterally until it divides into anteriorand posterior branches. Verify that the anterior branch occupies a deep groove or canal in the region of pterion. Look for the slender greater petrosal nerve passing posteromedial to the foramen spinosum and foramen ovale (Gr 7.22A, 7.23A) en route to foramen lacerum. We will study its origin and destination later. XXX
The structures in the posterior cranial fossa will be studied during the next lab.
The illustrations in this dissection guide are used with permission from Gray’s Anatomy for Students. 2005, by Richard Drake, Wayne Vogel, and Adam Mitchell, Elsevier Inc., Philadelphia; and from Grant’s Atlas of Anatomy, 11E, 2005, Anne Agur and Arthur Dalley II, Lippincott Williams & Wilkins, Philadelphia.